-
Fat Loss
-
Energy
-
Improved endurance
-
Bigger Muscles
-
Versatility
 Everybody who’s ever lifted weights wants to break their threshold for lifting, recovery and muscle growth. It’s never enough, is it?
Everybody who’s ever lifted weights wants to break their threshold for lifting, recovery and muscle growth. It’s never enough, is it?
But sadly, our bodies are not designed for bulked up biceps and tree trunk-thick thighs. So, after a while, it says, ‘You know what guys, that’s enough. That’s the best I can do.’
That’s when people start to look for help outside. Miracle pills and magic potions that can give them the body they desire without wreaking havoc with the endocrine, the central nervous and the reproductive systems. Sounds like a dream!
Cardarine, also known as Endurobol or GW1516, GW 501516, GW 501,516 and GSK-516 (That’s the last one. We promise) is as close to a magic potion as it can get. Did we sound blasphemous? The roid community is probably ripping their hair out as they read this. Oh wait, they lost all their hair already courtesy Winny!
Jokes apart, Cardarine is now one of the most widely used drugs in athletic sports and for good reason. It increases endurance, torches the fat that your body so loves to store and improves your insulin sensitivity which directly enhances protein synthesis.
To take it home, bigger muscles, faster recovery and fat loss.
So, what’s the catch?
Let’s take a closer look.
Table of Contents
What is Cardarine?
In layman’s terms, Cardaine is a drug that was formulated by GlaxoSmithKline (GSK) and Ligand Pharmaceuticals way back in 1992, for reducing the levels of LDL (bad) cholesterol and increasing HDL (good) cholesterol in the body. One of the purported uses was preventing and curing tumors formed in the breasts and the stomach.

First cycling positive for GW501516, Rusvelo’s Valery Kaykov suspended in 2013
As research continued in the early 2000s, it caught the eye of athletes and bodybuilders due to its ability to increase fatty acid metabolism. However, the effects, particularly boosting endurance, were so pronounced, that the drug was banned in professional sports. (Banned athletes for testing positive for Cardarine use: Valery Kaykov, Elena Lashmanova)
It’s not an Anabolic steroid mind you. But it can mimic the effect of exercise and amplify recovery, priming your body for better lifts, sooner than you can ever imagine.
Now it’s time for some scientific gobbledygook.
Cardarine is a PPARδ (Peroxisome Proliferator-Activator Receptor-delta) agonist that binds to the group of nuclear receptors in the body that oversee steroidal hormones. It increases gene expression and allows the body to expend energy even when it is at rest.
In simpler terms, you burn fat even when you are resting. If you are an athlete, say a marathoner or a bodybuilder, it’s like running on nitrous. You get an energy boost for a short amount of time allowing you to perform like never before. But, instead of catabolizing your hard earned muscle, your body now burns fat for energy.
How to use Cardarine
So, why isn’t everybody using it?
Despite all the positive press and the anecdotal instances that sound promising, there is very little clinical research on humans to warrant the long term use.
Most of the research has been conducted on mice.
So, you still have to cycle Cardarine, the way you’d cycle any SARM, AAS or other synthetic compound.
The average recommended dose is 10 mg/ml once a day. It has a half-life of about 24 hours. So you can either take all of it an hour before your workout or non-competitive sporting event or you can divide the dose into two for morning and evening.
Some bodybuilders also recommend upping the dose to 20 mg per day. That’s the upper limit for you.
Do not exceed this dose.
Authentic Cardarine is only available in liquid form and you can use the dropper to put the liquid in your mouth and gulp it down with juice or water.
A cycle can last from 4-6 weeks depending on your goals.
Whether or not you may need a PCT after a cycle of Cardarine is much debated. To play it safe, you can continue with a very basic Nolva-Clomid combo PCT for four weeks.
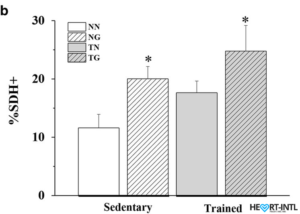
Study shows quantification of SDH-positive fibres in all groups (n = 3). *p < 0.05 compared to the NN group.
What can you expect after using Cardarine
That’s what it all boils down to. What can Cardarine do for you?
Here are some of the direct effects. We will speak about the indirect ones in a bit.
- Fat Loss: By activating PPARẟ, Cardarine amplifies the rate at which your body utilizes stored fat for energy. Insulin sensitivity is boosted and the output of glucose by the liver is reduced. You will look more ripped
- Energy: If you are on a cutting diet, your energy levels will not dip. If you lift, you will be able to break plateaus. If you run, get ready to breach your personal bests. Also, there’s no crash or jitters because it is not a stimulant. It gives you a consistent flow of energy that is manageable.
- Improved endurance: Cardarine increases the expression of the PPAR gene that is found in slow twitch muscle fibers which leads to increased oxygen use. You will recover faster and there will be very minimal muscle soreness.
- Bigger Muscles: A combination of the above factors allows you to lift better in the gym. It may not be anabolic but it hinders catabolism. Club that with a healthy diet with the right macronutrient ratios and you have all the right ingredients for muscle growth.
- Versatile: You can stack it with almost any compound without the risk or worry of side effects.
Is it safe for use?
Well, there haven’t been enough clinical studies on humans to be honest. And all the studies that have been conducted on rodents have been positive.
Having said that, athletes, bodybuilders and sportsmen who have been using it for the past 20 years swear by its effectiveness and tout it to be side effect free.
In all fairness, it must be mentioned that one clinical study with rodents showed an increase in the rate at which it caused colon cancer. But the doses that were used were outrageously high (1000 times the recommended dose) and for an extended time frame.
At normal doses, it would be safe to say that there are no known side effects.
Where can I buy authentic Cardarine?
By now, you are probably wondering where you can grab yourself a bottle of cardarine to gulp down. Hold your horses!
Like most other SARMS, there’s a high probability that the bottle of GW-501516 you buy may be fake. So, it’s always recommended that you buy from trusted sources only.
You can buy 100% pure and original cardarine here!




 Review){kind=link}