 Another day, another SARM (Selective Androgen Receptor Modulator) review on our hands.
Another day, another SARM (Selective Androgen Receptor Modulator) review on our hands.
Today, we look at MK-2866, fondly called Ostarine by fitness junkies.
Now, this is undoubtedly, one of the most widely used SARMs in the world. In fact, it is one of the first and only SARMs that has been clinically tested on humans. The other one of course, being Ligandrol (LGD 4033).
The tests are limited in number. But at least there’s irrefutable data that supports what has already been proven in multiple clinical tests involving mice. And then there’s your favorite bodybuilder or athlete who swears by the efficacy of the drug. That is if you are willing to take anecdotal evidence into consideration.
So, what is it about Ostarine that makes it so effective?
Let’s go analyze it.
Table of Contents
What is Ostarine (MK-2866)?
Ostarine, technically called MK-2866, GTx-024 and Enobosarm is a synthetic drug created by GTx INC, (based in Memphis) as an investigational selective androgen receptor modulator (SARM) for preventing muscle loss.
There were multiple clinical trials conducted for the purported use and it was found to be very effective in helping prevent dystrophy or muscle wasting caused due to cancer.
However, some of the results of the clinical trials were so pronounced that it introduced Ostarine into the bodybuilding domain.
Ever since, there has been tons of literature on the drug, usage and sides, albeit, most of it self-published.
But anyone who has used it calls it the safest and most anabolic SARM available for use. One that comes close to mimicking the effects of pure testosterone without interfering too much with your body’s natural test production.
It selectively boosts androgenic activity in muscle and bone tissue. Clubbed with a healthy diet and a rigorous exercise routine, that’s like being on a cycle of Test-E and probably even Dianabol without the nasties associated with them.
No androgenic activity in non-skeletal muscle tissues like the liver, the prostate or the brain.
How much of this is true though? Because if supplement manufacturers are to be believed, everything that they sell is anabolic and anti-catabolic without any side effects.
How to use Ostarine MK-2866
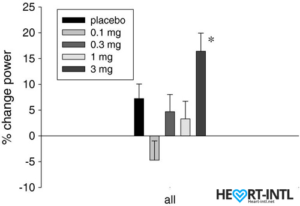
Stair climb power % change from baseline to day 86/End of Study
There are mixed reviews on the supposed usage. Some bodybuilders add it to an existing SARM stack as a pure bulking compound whereas others use it as a standalone compound for body re-composition.
It has an innate and unique ability to burn fat whilst increasing strength and lean muscular gains. Something that is almost considered impossible to achieve unless you are juicing.
While the recommended dosage is close to 15mg per day, most bodybuilders use up to 25 mg per day for a 12-week cycle.
Lean muscle gains of up to 7 lbs. are considered a normal result while using Ostarine. Whereas if you are on a controlled diet that is attuned to your goals, you should be able to achieve double-digit lean muscle mass gains in a cycle.
It has a half-life of 24 hours. So, one dose a day is all that is needed to keep the levels balanced in your blood stream.
What can you expect after using Ostarine?
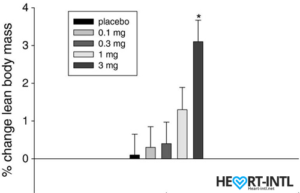
Lean Body Mass % change from baseline to day 86/End of Study
Depending on whether you are looking to cut or bulk, here are some of the typical results of using Ostarine.
- Bulking cycle: MK-2866 is an extremely anabolic synthetic drug. It will boost your strength, energy and reduce fatigue. More importantly, it boosts protein synthesis and will help you increase lean muscle mass. Notice how we mentioned ‘lean’? That’s because there is no water retention or puffiness that’s associated with steroids. Most importantly, the gains achieved with a cycle can be retained provided you continue with the same diet and exercise routine after cycle.
- Cutting cycle: While on a calorie deficit, your body can easily slip into a catabolic state created due to the lack of calories, a dented metabolism and a reduction in hormones like Insulin Growth Factor and Testosterone. MK-2866 prevents your hard earned muscle from being catabolized for fuel while you are in a deficit. This allows you to go into a cutting cycle without having to worry about muscle loss anymore. This is proven by at least two clinical studies. Off topic, S-4 is another SARM that is noted for its amazing effects during a cutting cycle.
- Body Recomposition: Considering that it allows you to retain muscle while you are cutting and gain lean muscle while you are bulking, staying a tad above your maintenance calories while on an MK2866 cycle allows you to recompose your body. You will lose fat and gain lean muscle. The gains won’t be as pronounced as it will be in a pure bulking cycle. But yes, you can easily gain 7-8 lbs. of lean muscle mass while losing at least a few inches off your waist.
Is it completely safe for use?
Like any other SARM, Ostarine is a non-hormonal synthetic drug. This means that it does not convert metabolically to an androgen. This also means that it will not aromatize into Estrogen.
So, no bitch tits.
However, blood work after a cycle of Ostarine has revealed a minimal increase in serum estradiol. It is a very mild increase mind you. But if you are concerned or have a high sensitivity to estrogen, then you can club it with mild or minimal doses of an Aromatize inhibitor like Arimidex.
Also, there will be mild testosterone suppression, especially at high doses of MK2866. So, if you are using it as a supplement, we highly recommend a three week PCT with Nolva or Clomid in small doses.
Where can I buy authentic MK2866?
Remember that the drug is not approved for use or sale as a supplement in the US. So, most of the products being peddled off as Ostarine is manufactured in China.
We wouldn’t trust any of them except for a trusted supplier that we have worked with and one that accepts credit card payments and not Bitcoin.
Here’s a link to our trusted supplier for SARMS and authentic MK2866.
-
Lean muscle Gain
-
Energy
-
Muscle retention




 Review){kind=link}